Visual cortex
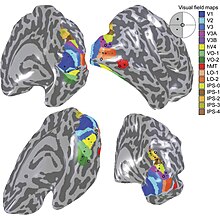
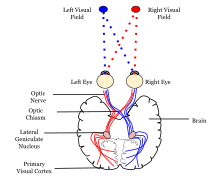
Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex.Other studies, however, provide strong support for the idea that skilled actions such as grasping are not affected by pictorial illusions[9][10] and suggest that the action/perception dissociation is a useful way to characterize the functional division of labor between the dorsal and ventral visual pathways in the cerebral cortex.Research on V1 has also revealed the presence of orientation-selective cells, which respond preferentially to stimuli with a specific orientation, contributing to the perception of edges and contours.Studies have demonstrated that sensory deprivation or exposure to enriched environments can lead to alterations in the organization and responsiveness of V1 neurons, highlighting the dynamic nature of this critical visual processing hub.The name "striate cortex" is derived from the line of Gennari, a distinctive stripe visible to the naked eye that represents myelinated axons from the lateral geniculate body terminating in layer 4 of the gray matter.In the case of the striate cortex, the line of Gennari corresponds to a band rich in myelinated nerve fibers, providing a clear marker for the primary visual processing region.It serves as a crucial hub for the initial processing of visual information, such as the analysis of basic features like orientation, spatial frequency, and color.Studies have revealed that changes in sensory input, such as those induced by visual training or deprivation, can lead to shifts in the retinotopic map.This adaptability underscores the brain's capacity to reorganize in response to varying environmental demands, highlighting the dynamic nature of visual processing.The correspondence between specific locations in V1 and the subjective visual field is exceptionally precise, even extending to map the blind spots of the retina.Notably, neurons in V1 have the smallest receptive field size, signifying the highest resolution, among visual cortex microscopic regions.Perhaps for the purpose of accurate spatial encoding, neurons in V1 have the smallest receptive field size (that is, the highest resolution) of any visual cortex microscopic regions.That is, the neuronal responses can discriminate small changes in visual orientations, spatial frequencies and colors (as in the optical system of a camera obscura, but projected onto retinal cells of the eye, which are clustered in density and fineness).David Hubel and Torsten Wiesel proposed the classic ice-cube organization model of cortical columns for two tuning properties: ocular dominance and orientation.V2 neurons build upon the basic features detected in V1, extracting more complex visual attributes such as texture, depth, and color.The responses of many V2 neurons are also modulated by more complex properties, such as the orientation of illusory contours,[33][34] binocular disparity,[35] and whether the stimulus is part of the figure or the ground.[36][37] Recent research has shown that V2 cells show a small amount of attentional modulation (more than V1, less than V4), are tuned for moderately complex patterns, and may be driven by multiple orientations at different subregions within a single receptive field.[38] This theory, unlike the dominant one, predicts that object-recognition memory (ORM) alterations could result from the manipulation in V2, an area that is highly interconnected within the ventral stream of visual cortices.[39][40] Most of the neurons of this area in primates are tuned to simple visual characteristics such as orientation, spatial frequency, size, color, and shape.The "complex" nomenclature is justified by the fact that some controversy still exists regarding the exact extent of area V3, with some researchers proposing that the cortex located in front of V2 may include two or three functional subdivisions.Neurons in area DM respond to coherent motion of large patterns covering extensive portions of the visual field (Lui and collaborators, 2006).[56] The pattern of projections to MT changes somewhat between the representations of the foveal and peripheral visual fields, with the latter receiving inputs from areas located in the midline cortex and retrosplenial region.[52] Nonetheless, several studies have demonstrated that neurons in MT are capable of responding to visual information, often in a direction-selective manner, even after V1 has been destroyed or inactivated.The first studies of the electrophysiological properties of neurons in MT showed that a large portion of the cells are tuned to the speed and direction of moving visual stimuli.[61] Neuropsychological studies of a patient unable to see motion, seeing the world in a series of static 'frames' instead, suggested that V5 in the primate is homologous to MT in the human.[65] There is still much controversy over the exact form of the computations carried out in area MT[66] and some research suggests that feature motion is in fact already available at lower levels of the visual system such as V1.[77][78] However, in comparison with area MT, a much smaller proportion of DM cells shows selectivity for the direction of motion of visual patterns.[79] Another notable difference with area MT is that cells in DM are attuned to low spatial frequency components of an image, and respond poorly to the motion of textured patterns such as a field of random dots.[80] The connections and response properties of cells in DM/V6 suggest that this area is a key node in a subset of the "dorsal stream", referred to by some as the "dorsomedial pathway".[81] This pathway is likely to be important for the control of skeletomotor activity, including postural reactions and reaching movements towards objects[76] The main 'feedforward' connection of DM is to the cortex immediately rostral to it, in the interface between the occipital and parietal lobes (V6A).



NeuroLexAnatomical terms of neuroanatomycerebral cortexvisual informationoccipital lobelateral geniculate nucleusthalamusBrodmann areaextrastriateBrodmann area 18Brodmann area 19hemispheres of the brainvisual fieldcalcarine fissureNeuronsaction potentialsreceptive fieldneuronal tuningretinotopiccalcarine branchposterior cerebral arteryTwo-streams hypothesisdorsal streamventral streaminferior temporal cortexlong-term memorydorsomedial areaposterior parietal cortexsaccadesUngerleiderMishkinGoodaleEbbinghaus illusionMicrographpia materarachnoid materblood vesselswhite matterHE-LFB stainpattern recognitionmyelinatedlateral geniculate bodygray mattermagnocellularparvocellularVisual systemblind spotscortical magnificationorientationsspatial frequenciescolorscamera obscuraretinalbinocular visioncortical columnsDavid HubelTorsten Wieselocular dominanceGabor transformfeedbackpyramidal neuronsedge detectionsaliency mapV1 Saliency Hypothesissuperior colliculusperception of illusionscolour centreAnatomical terminologydorsalventralhemispheresillusory contoursbinocular disparityparietal cortexglobal motionColor centermacaquesposterior inferotemporal area (PIT)homologuedorsal prelunate gyrusselective attentioninferotemporal cortexSemir Zekiprelunate gyrusfrontal eye fieldsNew World monkeysOld World monkeysperception of motioneye movementskoniocellularinferior pulvinarretrosplenial regionelectrophysiologicalLesionNeuropsychologicalJohn AllmanJon Kaasextrastriate cortexmedial longitudinal fissureparieto-occipital sulcuscommon marmosetprimary visual cortexmyelinnight monkeyscommon marmosetspremotor cortexCortical areaCortical blindnessFeature integration theoryList of regions in the human brainRetinotopyVisual processingComplex cellBibcodeSpringerCiteSeerXChurchland PSSejnowski TJThe Computational BrainMIT PressSaksida LMMoore TUniversity of UtahHarvard UniversityNeuroNamesBrainMaps project