Instead, the nucleoid forms by condensation and functional arrangement with the help of chromosomal architectural proteins and RNA molecules as well as DNA supercoiling.In E. coli, the chromosomal DNA is on average negatively supercoiled and folded into plectonemic loops, which are confined to different physical regions, and rarely diffuse into each other.In many bacteria, the chromosome is a single covalently closed (circular) double-stranded DNA molecule that encodes the genetic information in a haploid form.[54] A distinguishable feature of histone-like or heat-stable nucleoid structuring protein (H-NS)[55][56][57][58] from other NAPs is the ability to switch from the homodimeric form at relatively low concentrations (<1 x 10−5 M) to an oligomeric state at higher levels.[70][71] Taken together, it appears that the formation of rigid filaments is the most likely outcome of H-NS-DNA interactions in vivo that leads to gene silencing but does not induce DNA condensation.[74][75] Consistently, the crystal structure of Fis-DNA complexes shows that the distance between the recognition helices remains unchanged whereas DNA curves in the range of 60-75 degree.The loops in magnetic tweezers experiments are distinct from the micro-loops created by coherent DNA bending at cognate sites, as they require the formation of high-density DNA-protein complexes achieved by sequence-independent binding.The in-tandem occurrence of specific sites might initiate a nucleation reaction similar to that of H-NS, and then non-specific binding would lead to the formation of localized high-density Fis arrays.[47][83][84] Because of its helical structure, a double-stranded DNA molecule becomes topologically constrained in the covalently closed circular form which eliminates the rotation of the free ends.[91] Three factors contribute to generating and maintaining chromosomal DNA supercoiling in E. coli: (i) activities of topoisomerases, (ii) the act of transcription, and (iii) NAPs.[100][101] There is genetic evidence to suggest that a balance between the opposing activities of DNA gyrase and Topo I are responsible for maintaining a steady-state level of average negative superhelicity in E.It has been found that a topological constraint is not needed because RNAP generates sufficient torque that causes supercoiling even in a linear DNA template.[137][138] Specific topological changes by NAPs not only regulate gene transcription, but are also involved in other processes such as DNA replication initiation, recombination, and transposition.A positive correlation between DNA gyrase binding and upregulation of the genes caused by the absence of HU suggests that changes in supercoiling are responsible for differential expression.[143] These studies show that amino acid substitutions in HU can have a dramatic effect on nucleoid structure, that in turn results in significant phenotypic changes.[143] Since MukB and HU have emerged as critical players in long-range DNA interactions, it will be worthwhile to compare the effect of each of these two proteins on global gene expression.It appears that a supercoiling-diffusion barrier responsible for segregating plectonemic DNA loops into topological domains functions as a CID boundary in E. coli and many other bacteria.In E. coli, MDs were initially identified as large segments of the genome whose DNA markers localized together (co-localized) in fluorescence in situ hybridization (FISH) studies.[151][152] A large genomic region (~1-Mb) covering oriC (origin of chromosome replication) locus co-localized and was called Ori macrodomain.MDs were later identified based on how frequently pairs of lambda att sites that were inserted at various distant locations in the chromosome recombined with each other.[156] One obvious model based on in vitro results is that MatP promotes DNA-DNA contacts in vivo by bridging matS sites.Furthermore, in a strain already lacking MatP, the absence of MukB causes a reduction in DNA contacts throughout the chromosome, including the Ter domain.[126] It remains to be studied how DNA organization activities of Fis and HU that are well understood at a smaller scale (~1-kb) results in the formation of long-range DNA-DNA interactions.A recent study using fluorescent markers for detection of specific DNA loci examined pairwise physical distances between the seven rRNA operons that are genetically separated from each other (by as much as two million bp).[176][178] Conventional transmission electron microscopy (TEM) of chemically fixed E. coli cells portrayed the nucleoid as an irregularly shaped organelle.[13] A detailed examination of the 3D fluorescence image after cross-sectioning perpendicular to its long axis further revealed two global features of the nucleoid: curvature and longitudinal, high-density regions.Besides these examples, the chromosome is also specifically anchored to the cell membrane through protein-protein interaction between DNA-bound proteins, e.g., SlmA and MatP, and the divisome.The formation of highly condensed nucleoids upon the exposure of E. coli cells to chloramphenicol, which blocks translation, provides support for the expansion force of transient DNA-membrane contacts formed through transertion.Similar to B. subtilis and E. coli above, exposures of the archaean Haloferax volcanii to stresses that damage DNA cause compaction and reorganization of the nucleoid.[193] This article was adapted from the following source under a CC BY 4.0 license (2019) (reviewer reports): Subhash Verma; Zhong Qian; Sankar L Adhya (December 2019).
Formation of the
Escherichia coli
nucleoid
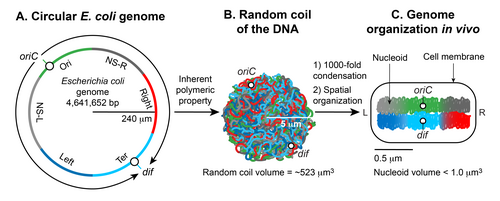
A.
An illustration of an open conformation of the circular genome of
Escherichia coli
. Arrows represent bi-directional DNA replication. The genetic position of the origin of bi-directional DNA replication (
oriC
) and the site of chromosome decatenation (
dif
) in the replication termination region (
ter
) are marked. Colors represent specific segments of DNA as discussed in C.
B.
An illustration of a random coil form adopted by the pure circular DNA of
Escherichia coli
at thermal equilibrium without supercoils and additional stabilizing factors.
[
4
]
[
5
]
C.
A cartoon of the chromosome of a newly born
Escherichia coli
cell. The genomic DNA is not only condensed by 1000-fold compared to its pure random coil form but is also spatially organized.
oriC
and
dif
are localized in the mid-cell, and specific regions of the DNA indicated by colors in A organize into spatially distinct domains.
Nucleoid at ≥1 kb scale. DNA organization by nucleoid-associated proteins.
DNA is depicted as a grey straight or curved line and the nucleoid-associated proteins are depicted as blue spheres.
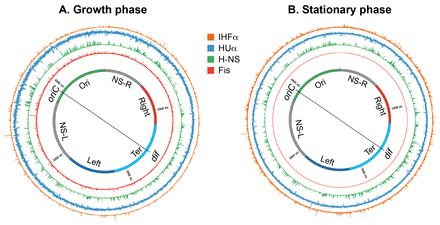
Genome-wide occupancy of nucleoid-associated proteins of
E. coli
.
A circular layout of the
E. coli
genome depicting genome-wide occupancy of NAPs Fis, H-NS, HU, and IHF in growth and stationary phases in
E. coli
. Histogram plots of the genome occupancy of NAPs as determined by chromatin-immunoprecipitation coupled with DNA sequencing (ChIP-seq) are shown outside the circular genome. The bin size of the histograms is 300 bp. Figure prepared in circos/0.69-6 using the ChIP-Seq data from.
[
46
]
[
50
]
DNA supercoiling
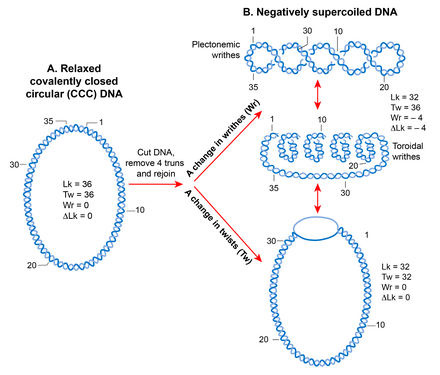
A.
A linear double-stranded DNA becomes a topologically constrained molecule if the two ends are covalently joined, forming a circle. Rules of DNA topology are explained using such a molecule (ccc-DNA) in which a numerical parameter called the linking number (Lk) defines the topology. Lk is a mathematical sum of two geometric parameters, twist (Tw) and writhe (Wr). A twist is the crossing of two strands, and writhe is coiling of the DNA double helix on its axis that requires bending. Lk is always an integer and remains invariant no matter how much the two strands are deformed. It can only be changed by introducing a break in one or both DNA strands by DNA metabolic enzymes called topoisomerases.
B.
A torsional strain created by a change in Lk of a relaxed, topologically constrained DNA manifests in the form of DNA supercoiling. A decrease in Lk (Lk<Lk
0
) induces negative supercoiling whereas an increase in Lk (Lk>Lk
0
) induces positive supercoiling. Only negative supercoiling is depicted here. For example, if a cut is introduced into a ccc-DNA and four turns are removed before rejoining the two strands, the DNA becomes negatively supercoiled with a decrease in the number of twists or writhe or both. Writhe can adopt two types of geometric structures called plectoneme and toroid. Plectonemes are characterized by the interwinding of the DNA double helix and an apical loop, whereas spiraling of DNA double helix around an axis forms toroids.
Basic units of genomic organization in bacteria and eukaryotes
Genomic DNA, depicted as a grey line, is negatively supercoiled in both bacteria and eukaryotes. However, the negatively supercoiled DNA is organized in the plectonemic form in bacteria, whereas it is organized in the toroidal form in eukaryotes. Nucleoid associated proteins (NAPs), shown as colored spheres, restrain half of the plectonemic supercoils, whereas almost all of the toroidal supercoils are induced as well as restrained by nucleosomes (colored orange), formed by wrapping of DNA around histones.
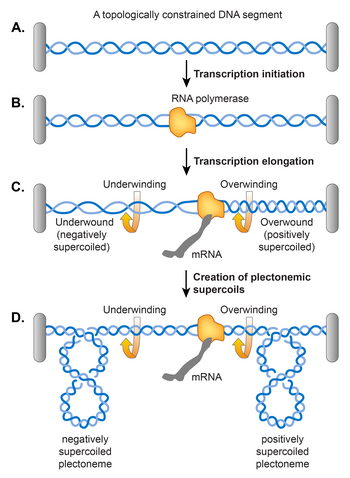
Twin supercoiling domain model for transcription-induced supercoiling
A.
An example of topologically constrained DNA. A grey bar represents a topological constraint, e.g. a protein or a membrane anchor.
B.
Accommodation of RNA polymerase for transcription initiation results in the opening of the DNA double helix.
C.
An elongating RNA polymerase complex cannot rotate around the helical axis of DNA. Therefore, removal of helical turns by RNA polymerase causes overwinding of the topologically constrained DNA ahead and underwinding of the DNA behind, generating positively and negatively supercoiled DNA, respectively. Supercoiling can manifest as either change in the numbers of twists as shown in C or plectonemic writhe as shown in D.
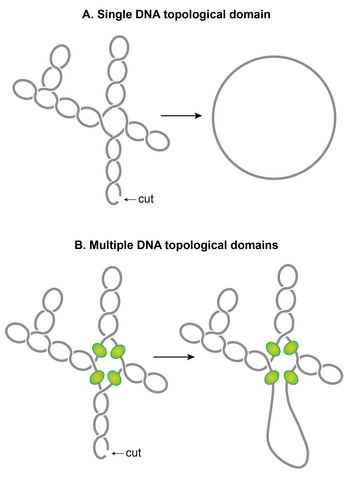
The chromosomal DNA within the nucleoid is segregated into independent supercoiled topological domains
A.
An illustration of a single topological domain of a supercoiled DNA. A single double-stranded cut anywhere would be sufficient to relax the supercoiling tension of the entire domain.
B.
An illustration of multiple topological domains in a supercoiled DNA molecule. A presence of supercoiling-diffusion barriers segregates a supercoiled DNA molecule into multiple topological domains. Hypothetical supercoiling diffusion barriers are represented as green spheres. As a result, a single double-stranded cut will only relax one topological domain and not the others. Plectonemic supercoils of DNA within the
E. coli
nucleoid are organized into several topological domains, but only four domains with a different number of supercoils are shown for simplicity.
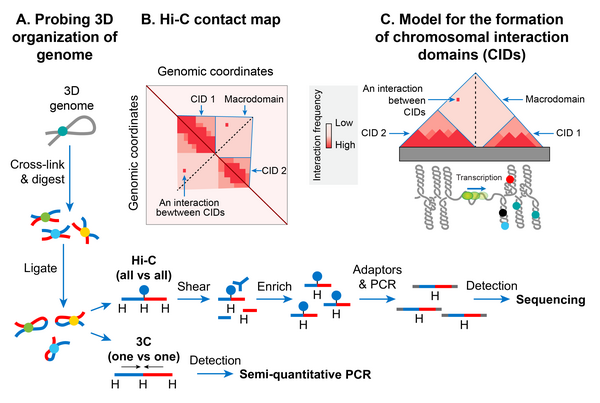
Nucleoid is spatially organized into chromosomal interactions domains (CIDs) and macrodomains
A.
Chromosome conformation capture (3C) methods probe 3D genome organization by quantifying physical interactions between genomic loci that are nearby in 3D-space but may be far away in the linear genome. A genome is cross-linked with formaldehyde to preserve physical contacts between genomic loci. Subsequently, the genome is digested with a restriction enzyme. In the next step, a DNA ligation is carried out under diluted DNA concentrations to favor intra-molecular ligation (between cross-linked fragments that are brought into physical proximity by 3D genome organization). A frequency of ligation events between distant DNA sites reflects a physical interaction. In the 3C method, ligation junctions are detected by the semi-quantitative PCR amplification in which amplification efficiency is a rough estimate of pairwise physical contact between genomic regions of interests and its frequency. The 3C method probes a physical interaction between two specific regions identified a priori, whereas its Hi-C version detects physical interactions between all possible pairs of genomic regions simultaneously. In the Hi-C method, digested ends are filled in with a biotinylated adaptor before ligation. Ligated fragments are sheared and then enriched by a biotin-pull down. Ligation junctions are then detected and quantified by the paired-end next-generation sequencing methods.
B.
Hi-C data are typically represented in the form of a two-dimensional matrix in which the x-axis and y-axis represent the genomic coordinates. The genome is usually divided into bins of a fixed size, e.g., 5-kb. The size of bins essentially defines the contact resolution. Each entry in the matrix, m
ij
, represents the number of chimeric sequencing reads mapped to genomic loci in bins i and j. A quantification of the reads (represented as a heatmap) denotes the relative frequency of contacts between genomic loci of bins i and j. A prominent feature of the heatmap is a diagonal line that appears due to more frequent physical interaction between loci that are very close to each other in the linear genome. The intensity further from the diagonal line represents the relative frequency of physical interaction between loci that are far away from each other in the linear genome. Triangles of high-intensity along the diagonal line represent highly self-interacting chromosomal interaction domains (CIDs) that are separated by a boundary region that consists of a smaller number of interactions.
C.
In many bacterial species including
E. coli
, it appears that supercoiled topological domains organize as CIDs. Plectonemic supercoiling promotes a high level of interaction among genomic loci within a CID, and a plectoneme-free region (PFR), created due to high transcription activity, acts as a CID boundary. Nucleoid-associated proteins, depicted as closed circles, stabilize the supercoiling-mediated interactions. The actively transcribing RNA polymerase (depicted as a green sphere) in the PFR blocks dissipation of supercoiling between the two domains thus acts as a supercoiling diffusion barrier. The size of the CIDs ranges between 30 and 400 kb. Several triangles (CIDs) merge to form a bigger triangle that represents a macrodomain. In other words, CIDs of a macrodomain physically interact with each other more frequently than with CIDs of a neighboring macrodomain or with genomic loci outside of that macrodomain. A macrodomain may comprise several CIDs. For simplicity, a macrodomain comprising only two CIDs is shown.
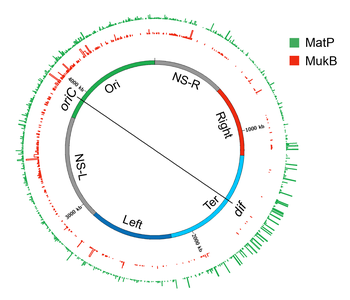
Genome-wide occupancy of MatP and MukB of
E. coli
A circular layout of the
E. coli
genome depicting genome-wide occupancy of MatP and MukB in
E. coli
. The innermost circle depicts the
E. coli
genome. The regions of the genome which organize as spatial domains(macrodomains) in the nucleoid are indicated as colored bands. Histogram plots of genome occupancy for MatP and MukB as determined by chromatin-immunoprecipitation coupled with DNA sequencing (ChIP-seq) are shown in outside circles. The bin size of the histograms is 300 bp. The figure was prepared in circos/0.69-6 using the processed ChIP-Seq data from.
[
155
]
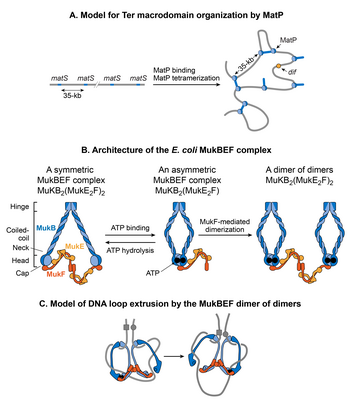
Models for DNA organization by MatP and MukBEF
A.
A
matS
-bridging model for DNA organization in the Ter macrodomain by MatP. MatP recognizes a 13-bp signature DNA sequence called
matS
that is present exclusively in the Ter macrodomain. There are 23
matS
sites separated by one another by an average of 35-kb. MatP binds to a
matS
site as a dimer, and the tetramerization of the DNA-bound dimers bridges
matS
sites forming large DNA loops.
B.
The architecture of the
E. coli
MukBEF complex. The complex is formed by protein-protein interactions between MukB (blue), MukF (dark orange) and MukE (light orange). MukB, which belongs to the family of structural maintenance of chromosomes (SMCs) proteins, forms a dimer (monomers are shown by dark and light blue colors) consisting of an ATPase head domain and a 100 nm long intramolecular coiled-coil with a hinge region in the middle. Because of the flexibility of the hinge region, MukB adopts a characteristic V-shape of the SMC family. MukF also tends to exist as a dimer because of the strong dimerization affinity between monomers.
[
157
]
[
158
]
The C-terminal domain of MukF can interact with the head domain of MukB while its central domain can interact with MukE. Two molecules of MukE and one molecule of MukF associate with each other independent of MukB to form a trimeric complex (MukE
2
F). Since MukF tends to exist in a dimeric form, the dimerization of MukF results in an elongated hexameric complex (MukE
2
F)
2
.
[
159
]
In the absence of ATP, the (MukE
2
F)
2
complex binds to the MukB head domains through the C-terminal domain of MukF to form a symmetric MukBEF complex (shown on the left). The stoichiometry of the symmetric complex is B
2
(E
2
F)
2
. The ATP binding between the MukB head domains forces the detachment of one MukF molecule and two MukE molecules.
[
132
]
[
159
]
As a result, an asymmetric MukBEF complex of the stoichiometry B
2
(E
2
F)
1
is formed. Since MukF readily dimerizes, the MukF dimerization can potentially join two ATP-bound asymmetric molecules resulting in the formation of a dimer of dimers with the stoichiometry of B
4
(E
2
F)
2
(shown on the right). The stoichiometry of the MukBEF complex
in vivo
is estimated to be B
4
(E
2
F)
2
suggesting that a dimer of dimers is the functional unit
in vivo
.
[
160
]
C.
A model for loop extrusion by a MukBEF dimer of dimers. A dimer of dimer loads onto DNA (depicted as a grey line) through DNA binding domains of MukB. MukB has been shown to bind DNA via its hinge region and the top region of its head domain.
[
48
]
[
161
]
The translocation of the complex away from its loading site then extrudes DNA loops. The loops are extruded in a rock-climbing manner by the coordinated opening and closing of the MukBEF ring through the MukB head disengagement that occurs due to coordinated ATP hydrolysis in the two dimers.
[
160
]
Dark and light blue circles represent ATP binding and hydrolysis events respectively. MukE is not shown in the complex for simplicity.
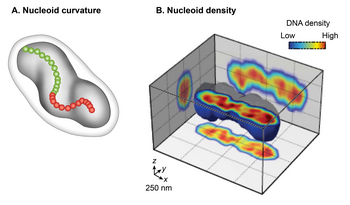
Nucleoid as a helical ellipsoid with longitudinal high-density DNA regions
A.
A cartoon of
E. coli
cell with a curved nucleoid (dark grey). A curved centroids path, denoted by red and green dots, emphasizes the curved shape of the nucleoid
[
13
]
B.
Cross-sectioning of the
E. coli
nucleoid visualized by HU-mCherry. Fluorescence intensity is taken as a proxy for DNA density and is represented by blue to red in increasing order.
[
14
]